Literature Background
Manoomin, wild rice, is an Indigenous cultural touchstone, food resource, and keystone wetland plant species found primarily in the Great Lakes region. Wild rice significantly influences wetland productivity, pollutant fate, and intricate relationships with various other plant and animal species. However, manoomin is rapidly disappearing across the heartland’s waters. Climate change, land development, and stripped protections threaten “the food that grows on water.” This page details the story of who manoomin is, its role in the environment, and the menagerie of its threats
Who is Manoomin?
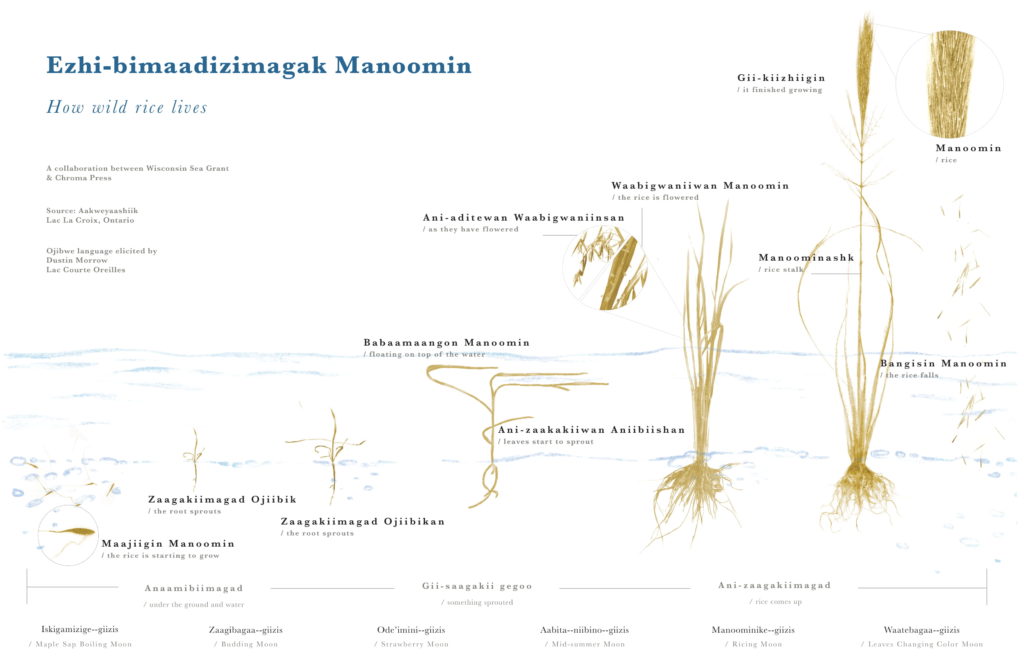
Wild rice is not actually a rice, but a grass. It is not related to white rice(Oryza) and differs from paddy wild rice grown and cultivated in parts of Minnesota and California. There are four species of natural wild rice but only two grow in the Midwest: Zizania Aquatica (Southern) and Zizania Palustris (Northern). Northern wild rice is adapted to colder climates. Southern wild rice is rarer and disappearing more rapidly.
Wild rice is a highly nutritious grain with significant benefits to cardiovascular health. While comparable to other grains, wild rice has a higher protein efficiency.
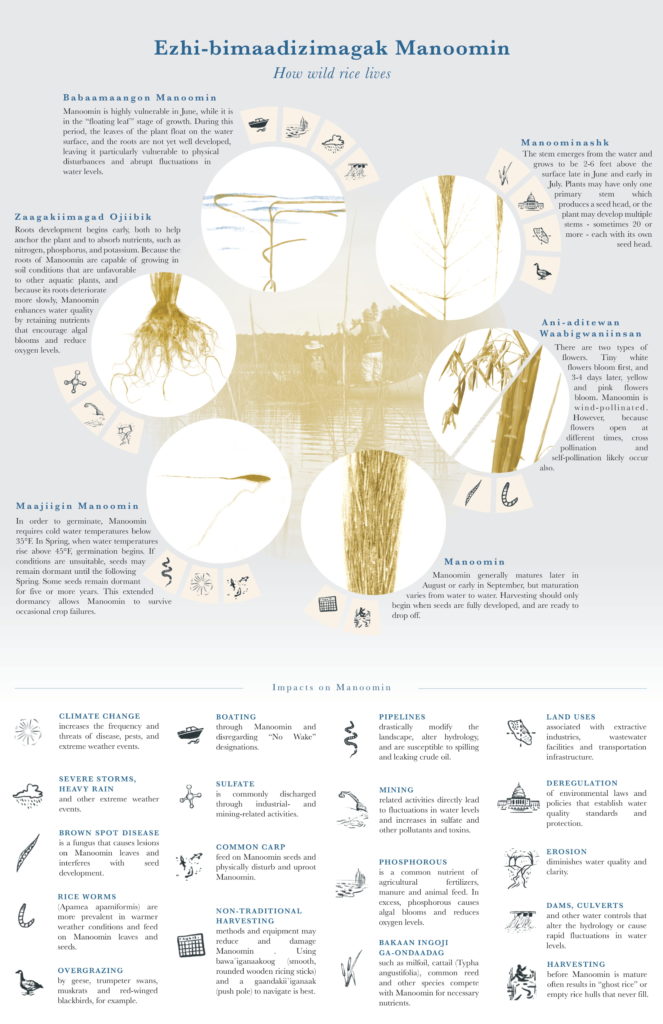
This resilient plant can survive in a variety of wetland conditions but thrives in 6inches to 3 feet water depths. Germinating in April, it grows for five months until reaching maturity.
At maturation in September, manoomin drops its seeds to embed in the sediment for dormancy until the next spring. This is when ricers collect rice by knocking them into canoes with ricing sticks or knockers. Harvesting is an important part of the self-sustaining nature of these environments.
Ecological Importance
Wild rice has a highly specialized role in the region’s wetlands because it is virtually the only native annual aquatic macrophyte in the Great Lakes region. During its growing season(April through September), manoomin beds can be nurseries, habitats, and food for a diverse range of animals including invertebrates, fish, muskrat, waterfowl, moose, and others. Although the annual plant completes its life cycle in one growing season and dies in the fall, the plant matter provides food and shelter to small aquatic animals year-round.
Manoomin also plays a critical role in water quality. Wild rice roots can safely remove and retain harmful chemicals from the environment (e.g., lead and arsenic) and efficiently absorb nutrients (e.g., nitrogen and phosphorus). The uptake of nutrients lowers nutrient availability to algae, potentially preventing harmful algal blooms and eutrophication.
Additionally, manoomin beds stabilize wetland sediments by preventing shoreline erosion and slowing the speed of flood waters. Seeds dropped in the fall can stay dormant for two to five years. However, the Anishinaabe have shared stories of wild rice staying dormant for decades until water quality improved.
Declines and Threats
Manoomin populations have dramatically declined between the 1900 and 2010 across the Great Lakes region due to both human-caused and natural disturbances. Recent observations indicate steeper population declines since 2010. A warming climate, land development, and unbalanced relationships weave an intricate tapestry of challenges to manoomin survival.
Physical disturbance is a significant threat to all life stages. As a warming climate increases storm frequency and severity, fast moving waters during the floating leaf stage can knock over and kill rice. Boat wake knocking over rice in June remains a significant threat despite various efforts to educate boaters.
Environmental shifts from climate change may be more favorable for competing, hardier (often invasive) species like Eurasian water milfoil and natural and hybrid cattails. Shorter and warmer winters prevent seeds from germinating the next year because seeds need at least 3 months under freezing or near freezing water conditions to successfully germinate in the spring.
Poor water quality also contributes to manoomin declines. Sulfides from mining operations are directly toxic to wild rice seedlings and can prevent mature rice from absorbing nitrogen later in its lifecycle. Water clarity is critical for early stages to access sunlight and oxygen.
Humans can also directly impact wild rice. Some people rip rice plants out to access waterways more easily for boating and recreation. Harvesting rice before it is ripe can prevent viable seeds from forming and dropping for the next season.
Finally, manoomin is threatened by disease (i.e., brown spot disease) and pests. Rice worms, which are thriving due to warmer summers and winters, can reduce manoomin seed production. Birds and fish, like common carp, can also overgraze wild rice. Carp also lay eggs in wild rice beds that when hatched, dishevel early rice life stages.
The extensive ecological functions and threats to “The Good Berry” illustrate a story of the hardship, resilience, and adaptability of wild rice.
References
Definition of keystone species: https://www.britannica.com/science/keystone-species
GWOW Website overview: http://www.g-wow.org/en-us/wildrice/default.aspx
Myrbo, A., Swain, E. B., Johnson, N. W., Engstrom, D. R., Pastor, J., Dewey, B., Monson, P., Brenner, J., Dykhuizen Shore, M., & Peters, E. B. (2017). Increase in Nutrients, Mercury, and Methylmercury as a Consequence of Elevated Sulfate Reduction to Sulfide in Experimental Wetland Mesocosms. Journal of Geophysical Research: Biogeosciences, 122(11), 2769–2785. https://doi.org/10.1002/2017JG003788
Pollman, C. D., Swain, E. B., Bael, D., Myrbo, A., Monson, P., & Shore, M. D. (2017). The Evolution of Sulfide in Shallow Aquatic Ecosystem Sediments: An Analysis of the Roles of Sulfate, Organic Carbon, and Iron and Feedback Constraints Using Structural Equation Modeling. Journal of Geophysical Research: Biogeosciences, 122(11), 2719–2735. https://doi.org/10.1002/2017JG003785
Pastor, J., Dewey, B., Johnson, N. W., Swain, E. B., Monson, P., Peters, E. B., & Myrbo, A. (2016). Effects of sulfate and sulfide on the life cycle of Zizania palustris in hydroponic and mesocosm experiments. Ecological Applications, 27(1), 321–336. https://doi.org/10.1002/eap.1452
Pastor, J., Walker, R. D., & Larsson, S. (2006). Delays in Nutrient Cycling and Plant Population Oscillations. Oikos, 112(3), 698–705. http://www.jstor.org.ezproxy.library.wisc.edu/stable/3548736
Walker, R. E. D., Pastor, J., & Dewey, B. W. (2010). Litter Quantity and Nitrogen Immobilization Cause Oscillations in Productivity of Wild Rice (Zizania palustris L.) in Northern Minnesota. Ecosystems, 13(4), 485–498. http://www.jstor.org.ezproxy.library.wisc.edu/stable/40793004
Yost, C. L., Blinnikov, M. S., & Julius, M. L. (2013). Detecting ancient wild rice (Zizania spp. L.) using phytoliths: a taphonomic study of modern wild rice in Minnesota (USA) lake sediments. Journal of Paleolimnology, 49(2), 221–236. https://doi.org/10.1007/s10933-012-9670-x
Jorgenson, K. D., Lee, P. F., & Kanavillil, N. (2012). Ecological relationships of wild rice, Zizania spp. 11. Electron microscopy study of iron plaques on the roots of northern wild rice (Zizania palustris). Botany, 91(3), 189–201. https://doi.org/10.1139/cjb-2012-0198
Hildebrandt, L. R., Pastor, J., & Dewey, B. (2012). Effects of external and internal nutrient supplies on decomposition of wild rice, Zizania palustris. Aquatic Botany, 97(1), 35–43. https://doi.org/https://doi.org/10.1016/j.aquabot.2011.11.002
Fort, D. J., Mathis, M. B., Walker, R., Tuominen, L. K., Hansel, M., Hall, S., Richards, R., Grattan, S. R., & Anderson, K. (2014). Toxicity of sulfate and chloride to early life stages of wild rice (Zizania palustris). Environmental Toxicology and Chemistry, 33(12), 2802–2809. https://doi.org/10.1002/etc.2744
Durkee Walker, R., Pastor, J., & Dewey, B. W. (2006). Effects of wild rice (Zizania palustris) straw on biomass and seed production in northern Minnesota. Canadian Journal of Botany, 84(6), 1019–1024. https://doi.org/10.1139/b06-058
Sims, L., Pastor, J., Lee, T., & Dewey, B. (2012). Nitrogen, phosphorus and light effects on growth and allocation of biomass and nutrients in wild rice. Oecologia, 170(1), 65–76. http://www.jstor.org.ezproxy.library.wisc.edu/stable/23259751
Sims, L., Pastor, J., Lee, T., & Dewey, B. (2012). Nitrogen, phosphorus, and light effects on reproduction and fitness of wild rice. Botany, 90(9), 876–883. https://doi.org/10.1139/b2012-057
Aagaard, K., Eash, J., Ford, W., Heglund, P. J., McDowell, M., & Thogmartin, W. E. (2018). Modeling the Relationship between Water Level, Wild Rice Abundance, and Waterfowl Abundance at a Central North American Wetland. Wetlands. https://doi.org/10.1007/s13157-018-1025-6
Dahlberg, Nathan B. and Pastor, J. (2014). Desirable Host Plant Qualities in Wild Rice (Zizania Palustris)for Infestation by the Rice Worm Apamea Apamiformis(Lepidoptera: Noctuidae). The Great Lakes Entomologist, 47(1), Article 6.
Myrbo, A., Swain, E. B., Engstrom, D. R., Coleman Wasik, J., Brenner, J., Dykhuizen Shore, M., Peters, E. B., & Blaha, G. (2017). Sulfide Generated by Sulfate Reduction is a Primary Controller of the Occurrence of Wild Rice (Zizania palustris) in Shallow Aquatic Ecosystems. Journal of Geophysical Research: Biogeosciences, 122(11), 2736–2753. https://doi.org/10.1002/2017JG003787
Surendiran, G., Alsaif, M., Kapourchali, F. R., & Moghadasian, M. H. (2014). Nutritional constituents and health benefits of wild rice (Zizania spp.). Nutrition Reviews, 72(4), 227–236. http://dx.doi.org/10.1111/nure.12101